
Page 12 - Demo
P. 12
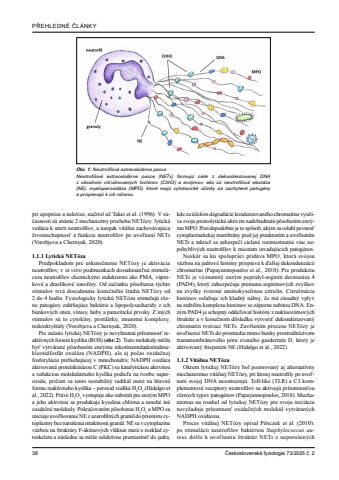
38 %u010ceskoslovensk%u00e1 fyziologie 73/2025 %u010d. 2P%u0158EHLEDN%u00c9 %u010cL%u00c1NKYpri apopt%u00f3ze a nekr%u00f3ze, na%u010drtol u%u017e Takei et al. (1996). V s%u00fa%u010dasnosti s%u00fa zn%u00e1me 2 mechanizmy priebehu NET%u00f3zy: lytick%u00e1 ved%u00faca k smrti neutrofilov, a naopak vit%u00e1lna zachov%u00e1vaj%u00faca %u017eivotaschopnos%u0165 a funkcie neutrofilov po uvo%u013enen%u00ed NETs (Vorobjeva a Chernyak, 2020).1.1.1 Lytick%u00e1 NET%u00f3zaPredpokladom pre uskuto%u010dnenie NET%u00f3zy je aktiv%u00e1cia neutrofilov, v in vitro podmienkach dosiahnute%u013en%u00e1 stimul%u00e1ciou neutrofilov chemick%u00fdmi induktormi ako PMA, v%u00e1pnikov%u00e9 a drasl%u00edkov%u00e9 ionof%u00f3ry. Od za%u010diatku p%u00f4sobenia t%u00fdchto stimulov trv%u00e1 dosiahnutie kone%u010dn%u00e9ho %u0161t%u00e1dia NET%u00f3zy od2 do 4 hod%u00edn. Fyziologicky lytick%u00fa NET%u00f3zu stimuluj%u00fa r%u00f4zne patog%u00e9ny zah%u0155%u0148aj%u00face bakt%u00e9rie a lipopolysacharidy z ich bunkov%u00fdch stien, v%u00edrusy, huby a parazitick%u00e9 prvoky. Z in%u00fdch stimulov s%u00fa to cytok%u00edny, protil%u00e1tky, imunitn%u00e9 komplexy, mikrokry%u0161t%u00e1ly (Vorobjeva a Chernyak, 2020).Pre za%u010datie lytickej NET%u00f3zy je nevyhnutn%u00e1 pr%u00edtomnos%u0165 reakt%u00edvnych foriem kysl%u00edka (ROS) (obr. 2). Tieto molekuly m%u00f4%u017eu by%u0165 vytv%u00e1ran%u00e9 p%u00f4soben%u00edm enz%u00fdmu nikot%u00ednamidaden%u00edndinukleotidfosf%u00e1t oxid%u00e1za (NADPH), ale aj po%u010das oxida%u010dnej fosforyl%u00e1cie prebiehaj%u00facej v mitochondrii; NADPH oxid%u00e1za aktivovan%u00e1 prote%u00ednkin%u00e1zou C (PKC) sa katalytickou aktivitou a redukciou molekul%u00e1rneho kysl%u00edka podie%u013ea na tvorbe superoxidu, pri%u010dom sa tento nestabiln%u00fd radik%u00e1l men%u00ed na hlavn%u00fa formu reakt%u00edvneho kysl%u00edka %u2013 peroxid vod%u00edka H2O2 (Hidalgo et al., 2022). Pr%u00e1ve H2O2 vystupuje ako substr%u00e1t pre enz%u00fdm MPO a jeho aktivitou sa produkuje kyselina chl%u00f3rna a mnoh%u00e9 in%u00e9 oxida%u010dn%u00e9 molekuly. Pokra%u010dovan%u00edm p%u00f4sobenia H2O2 a MPO sa iniciuje uvo%u013e%u0148ovanie NE z azurofiln%u00fdch gran%u00fal do priestoru cytoplazmy bez naru%u0161enia intaktnosti gran%u00fal. NE sa v cytoplazme v%u00e4zbou na %u0161trukt%u00fary F-akt%u00ednov%u00fdch vl%u00e1kien star%u00e1 o rozklad cytoskeletu a n%u00e1sledne sa m%u00f4%u017ee selekt%u00edvne premiestni%u0165 do jadra, kde za %u00fa%u010delom degrad%u00e1cie kondenzovan%u00e9ho chromat%u00ednu vyu%u017e%u00edva svoju proteolytick%u00fa aktivitu nadobudnut%u00fa p%u00f4soben%u00edm enz%u00fdmu MPO. Pravdepodobne je to sp%u00f4sob, ak%u00fdm sa oslab%u00ed pevnos%u0165 cytoplazmatickej membr%u00e1ny pred jej prasknut%u00edm a uvo%u013enen%u00edm NETs a taktie%u017e sa zabezpe%u010d%u00ed cielen%u00e9 rozmiestnenie viac nepohybliv%u00fdch neutrofilov k miestam invaduj%u00facich patog%u00e9nov.Nesk%u00f4r sa ku spolupr%u00e1ci prid%u00e1va MPO, ktor%u00e1 svojou v%u00e4zbou na jadrov%u00e9 hist%u00f3ny prispieva k %u010fal%u0161ej dekondenz%u00e1cii chromat%u00ednu (Papayannopoulos et al., 2010). Pre produkciu NETs je v%u00fdznamn%u00fd enz%u00fdm peptidyl-argin%u00edn deimin%u00e1za 4 (PAD4), ktor%u00fd zabezpe%u010duje premenu argin%u00ednov%u00fdch zvy%u0161kov na zvy%u0161ky tvoren%u00e9 aminokyselinou citrul%u00edn. Citrulin%u00e1cia hist%u00f3nov oslabuje ich kladn%u00fd n%u00e1boj, %u010do m%u00e1 z%u00e1sadn%u00fd vplyv na stabilitu komplexu hist%u00f3nov so z%u00e1porne nabitou DNA. Enz%u00fdm PAD4 je schopn%u00fd odde%u013eova%u0165 hist%u00f3ny z nukleoz%u00f3mov%u00fdch %u0161trukt%u00far a v kone%u010dnom d%u00f4sledku vytvori%u0165 dekondenzovan%u00fd chromat%u00edn tvoriaci NETs. Zav%u0155%u0161en%u00edm procesu NET%u00f3zy je uvo%u013enenie NETs do prostredia mimo bunky prostredn%u00edctvom transmembr%u00e1nov%u00e9ho p%u00f3ru zvan%u00e9ho gasderm%u00edn D, ktor%u00fd je aktivovan%u00fd %u0161tiepen%u00edm NE (Hidalgo et al., 2022).1.1.2 Vit%u00e1lna NET%u00f3zaOkrem lytickej NET%u00f3zy bol pozorovan%u00fd aj alternat%u00edvny mechanizmus vit%u00e1lnej NET%u00f3zy, pri ktorej neutrofily po uvo%u013enen%u00ed svojej DNA nezomieraj%u00fa. Toll-like (TLR) a C3 komplementov%u00e9 receptory neutrofilov sa aktivuj%u00fa pr%u00edtomnos%u0165ou r%u00f4znych typov patog%u00e9nov (Papayannopoulos, 2018). Mechanizmus na rozdiel od lytickej NET%u00f3zy pre svoju inici%u00e1ciu nevy%u017eaduje pr%u00edtomnos%u0165 oxida%u010dn%u00fdch molek%u00fal vytv%u00e1ran%u00fdch NADPH oxid%u00e1zou. Proces vit%u00e1lnej NET%u00f3zy op%u00edsal Pilsczek et al. (2010): po stimul%u00e1cii neutrofilov bakt%u00e9riou Staphylococcus aureus do%u0161lo k uvo%u013eneniu %u0161trukt%u00far NETs z neporu%u0161en%u00fdch Obr. 1: Neutrofilov%u00e1 extracelul%u00e1rna pascaNeutrofilov%u00e9 extracelul%u00e1rne pasce (NETs) formuj%u00fa siete z dekondenzovanej DNA s obsahom citrulinovan%u00fdch hist%u00f3nov (CitH3) a enz%u00fdmov, ako s%u00fa neutrofilov%u00e1 elast%u00e1za (NE), myeloperoxid%u00e1za (MPO), ktor%u00e9 maj%u00fa cytotoxick%u00e9 %u00fa%u010dinky na zachyten%u00e9 patog%u00e9ny a prispievaj%u00fa k ich ni%u010deniu.
