
Page 15 - YORAM RUDY BOOK FINAL
P. 15
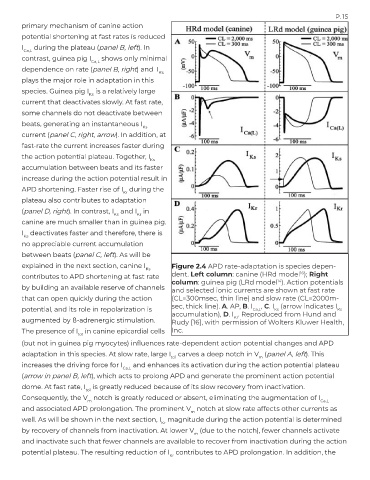
P. 15
primary mechanism of canine action
potential shortening at fast rates is reduced
I during the plateau (panel B, left). In
Ca,L
contrast, guinea pig I shows only minimal
Ca,L
dependence on rate (panel B, right) and I
Ks
plays the major role in adaptation in this
species. Guinea pig I is a relatively large
Ks
current that deactivates slowly. At fast rate,
some channels do not deactivate between
beats, generating an instantaneous I
Ks
current (panel C, right, arrow). In addition, at
fast-rate the current increases faster during
the action potential plateau. Together, I
Ks
accumulation between beats and its faster
increase during the action potential result in
APD shortening. Faster rise of I during the
Kr
plateau also contributes to adaptation
(panel D, right). In contrast, I and I in
Ks Kr
canine are much smaller than in guinea pig.
I deactivates faster and therefore, there is
Ks
no appreciable current accumulation
between beats (panel C, left). As will be
explained in the next section, canine I Figure 2.4 APD rate-adaptation is species depen-
Ks
16
contributes to APD shortening at fast rate dent. Left column: canine (HRd model ); Right
column: guinea pig (LRd model ). Action potentials
14
by building an available reserve of channels and selected ionic currents are shown at fast rate
that can open quickly during the action (CL=300msec, thin line) and slow rate (CL=2000m-
potential, and its role in repolarization is sec, thick line). A. AP, B. I Ca,L , C. I (arrow indicates I
Ks
Ks
accumulation), D. I . Reproduced from Hund and
Kr
augmented by ß-adrenergic stimulation. Rudy [16], with permission of Wolters Kluwer Health,
The presence of I in canine epicardial cells Inc.
to1
(but not in guinea pig myocytes) influences rate-dependent action potential changes and APD
adaptation in this species. At slow rate, large I carves a deep notch in V (panel A, left). This
to1 m
increases the driving force for I and enhances its activation during the action potential plateau
Ca,L
(arrow in panel B, left), which acts to prolong APD and generate the prominent action potential
dome. At fast rate, I is greatly reduced because of its slow recovery from inactivation.
to1
Consequently, the V notch is greatly reduced or absent, eliminating the augmentation of I
m Ca,L
and associated APD prolongation. The prominent V notch at slow rate affects other currents as
m
well. As will be shown in the next section, I magnitude during the action potential is determined
Kr
by recovery of channels from inactivation. At lower V (due to the notch), fewer channels activate
m
and inactivate such that fewer channels are available to recover from inactivation during the action
potential plateau. The resulting reduction of I contributes to APD prolongation. In addition, the
Kr