
Page 425 - The Toxicology of Fishes
P. 425
The Osmoregulatory System 405
chloride cell number
–2
(n.mm )
250
200
150
100
50
5 4 5 6
0
control 50 Cu 100 Cu 200 Cu
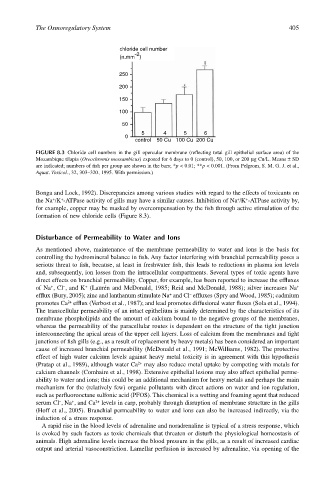
FIGURE 8.3 Chloride cell numbers in the gill opercular membrane (reflecting total gill epithelial surface area) of the
Mozambique tilapia (Oreochromis mossambicus) exposed for 6 days to 0 (control), 50, 100, or 200 µg Cu/L. Means ± SD
are indicated; numbers of fish per group are shown in the bars; *p < 0.01; **p < 0.001. (From Pelgrom, S. M. G. J. et al.,
Aquat. Toxicol., 32, 303–320, 1995. With permission.)
Bonga and Lock, 1992). Discrepancies among various studies with regard to the effects of toxicants on
+
the Na /K -ATPase activity of gills may have a similar causes. Inhibition of Na /K -ATPase activity by,
+
+
+
for example, copper may be masked by overcompensation by the fish through active stimulation of the
formation of new chloride cells (Figure 8.3).
Disturbance of Permeability to Water and Ions
As mentioned above, maintenance of the membrane permeability to water and ions is the basis for
controlling the hydromineral balance in fish. Any factor interfering with branchial permeability poses a
serious threat to fish, because, at least in freshwater fish, this leads to reductions in plasma ion levels
and, subsequently, ion losses from the intracellular compartments. Several types of toxic agents have
direct effects on branchial permeability. Copper, for example, has been reported to increase the effluxes
+
+
–
of Na , Cl , and K (Laurén and McDonald, 1985; Reid and McDonald, 1988); silver increases Na +
efflux (Bury, 2005); zinc and lanthanum stimulate Na and Cl effluxes (Spry and Wood, 1985); cadmium
–
+
2+
promotes Ca efflux (Verbost et al., 1987); and lead promotes diffusional water fluxes (Sola et al., 1994).
The transcellular permeability of an intact epithelium is mainly determined by the characteristics of its
membrane phospholipids and the amount of calcium bound to the negative groups of the membranes,
whereas the permeability of the paracellular routes is dependent on the structure of the tight junction
interconnecting the apical areas of the upper cell layers. Loss of calcium from the membranes and tight
junctions of fish gills (e.g., as a result of replacement by heavy metals) has been considered an important
cause of increased branchial permeability (McDonald et al., 1991; McWilliams, 1982). The protective
effect of high water calcium levels against heavy metal toxicity is in agreement with this hypothesis
(Pratap et al., 1989), although water Ca may also reduce metal uptake by competing with metals for
2+
calcium channels (Comhaire et al., 1998). Extensive epithelial lesions may also affect epithelial perme-
ability to water and ions; this could be an additional mechanism for heavy metals and perhaps the main
mechanism for the (relatively few) organic pollutants with direct actions on water and ion regulation,
such as perfluorooctane sulfonic acid (PFOS). This chemical is a wetting and foaming agent that reduced
serum Cl , Na , and Ca levels in carp, probably through disruption of membrane structure in the gills
2+
+
–
(Hoff et al., 2005). Branchial permeability to water and ions can also be increased indirectly, via the
induction of a stress response.
A rapid rise in the blood levels of adrenaline and noradrenaline is typical of a stress response, which
is evoked by such factors as toxic chemicals that threaten or disturb the physiological homeostasis of
animals. High adrenaline levels increase the blood pressure in the gills, as a result of increased cardiac
output and arterial vasoconstriction. Lamellar perfusion is increased by adrenaline, via opening of the