
Page 429 - Small Animal Clinical Nutrition 5th Edition
P. 429
Enteral-Assisted Feeding 443
Patients undergoing food deprivation display almost com-
VetBooks.ir plete reversal of the metabolic processes described for the well-
fed patient. Due to the lack of exogenous dietary sources,
endogenous sources become the primary fuel for meeting
immediate metabolic needs. Glycogen stores, instead of being
replenished, become exhausted as the initial energy source.
Then in order to preserve vital functions as long as possible,
patients use different proportions of stored body fat and protein
to maintain blood glucose concentrations. Which fuel or mix-
ture of fuels the patient uses depends on the length of time the
patient was food deprived and the quantity of each of the fuel
stores available to the patient (Figure 25-1). The adaptation
from fed to starved state is one in which fuel use by the patient
shifts from primarily a mixture of fuels to one in which the pri-
mary fuel is fatty acids.
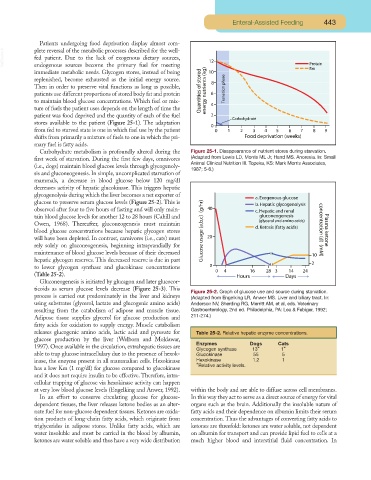
Carbohydrate metabolism is profoundly altered during the Figure 25-1. Disappearance of nutrient stores during starvation.
first week of starvation. During the first few days, omnivores (Adapted from Lewis LD, Morris ML Jr, Hand MS. Anorexia. In: Small
Animal Clinical Nutrition III. Topeka, KS: Mark Morris Associates,
(i.e., dogs) maintain blood glucose levels through glycogenoly-
1987; 5-6.)
sis and gluconeogenesis. In simple, uncomplicated starvation of
mammals, a decrease in blood glucose below 120 mg/dl
decreases activity of hepatic glucokinase. This triggers hepatic
glycogenolysis during which the liver becomes a net exporter of
glucose to preserve serum glucose levels (Figure 25-2). This is
observed after four to five hours of fasting and will only main-
tain blood glucose levels for another 12 to 28 hours (Cahill and
Owen, 1968). Thereafter, gluconeogenesis must maintain
blood glucose concentrations because hepatic glycogen stores
will have been depleted. In contrast, carnivores (i.e., cats) must
rely solely on gluconeogenesis, beginning intraprandially for
maintenance of blood glucose levels because of their decreased
hepatic glycogen reserves. This decreased reserve is due in part
to lower glycogen synthase and glucokinase concentrations
(Table 25-2).
Gluconeogenesis is initiated by glucagon and later glucocor-
ticoids as serum glucose levels decrease (Figure 25-3). This
Figure 25-2. Graph of glucose use and source during starvation.
process is carried out predominately in the liver and kidneys (Adapted from Engelking LR, Anwer MS. Liver and biliary tract. In:
using substrates (glycerol, lactate and glucogenic amino acids) Anderson NV, Sherding RG, Merritt AM, et al, eds. Veterinary
resulting from the catabolism of adipose and muscle tissue. Gastroenterology, 2nd ed. Philadelphia, PA: Lea & Febiger, 1992;
Adipose tissue supplies glycerol for glucose production and 211-274.)
fatty acids for oxidation to supply energy. Muscle catabolism
releases glucogenic amino acids, lactic acid and pyruvate for Table 25-2. Relative hepatic enzyme concentrations.
glucose production by the liver (Welborn and Moldawar,
Enzymes Dogs Cats
1997). Once available in the circulation, extrahepatic tissues are
Glycogen synthase 13* 1*
able to trap glucose intracellulary due to the presence of hexok- Glucokinase 55 5
inase, the enzyme present in all mammalian cells. Hexokinase Hexokinase 1.2 1
*Relative activity levels.
has a low Km (1 mg/dl) for glucose compared to glucokinase
and it does not require insulin to be effective. Therefore, intra-
cellular trapping of glucose via hexokinase activity can happen
at very low blood glucose levels (Engelking and Anwer, 1992). within the body and are able to diffuse across cell membranes.
In an effort to conserve circulating glucose for glucose- In this way they act to serve as a direct source of energy for vital
dependent tissues, the liver releases ketone bodies as an alter- organs such as the brain. Additionally the insoluble nature of
nate fuel for non-glucose dependent tissues. Ketones are oxida- fatty acids and their dependence on albumin limits their serum
tion products of long-chain fatty acids, which originate from concentration. Thus the advantages of converting fatty acids to
triglycerides in adipose stores. Unlike fatty acids, which are ketones are threefold: ketones are water soluble, not dependent
water insoluble and must be carried in the blood by albumin, on albumin for transport and can provide lipid fuel to cells at a
ketones are water soluble and thus have a very wide distribution much higher blood and interstitial fluid concentration. In